МУЖСКИЕ ПОЛОВЫЕ ОРГАНЫ (organa genitalia masculine)
В состав мужских половых органов, входят: яички с их оболочками, семявыносящие протоки с семенными пузырьками, предстательная железа, бульбоуретральные железы, половой член, состоящий из пещеристых тел. Здесь также будет описан и мужской мочеиспускательный канал, носящий смешанный характер мочеполовой трубки.
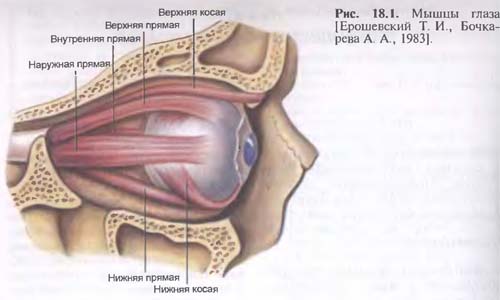
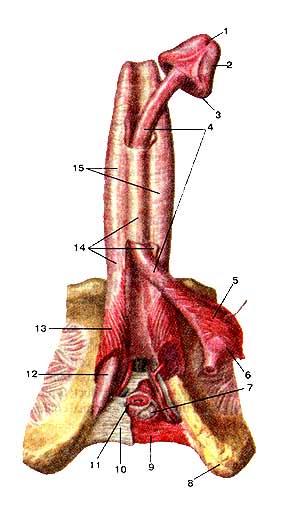
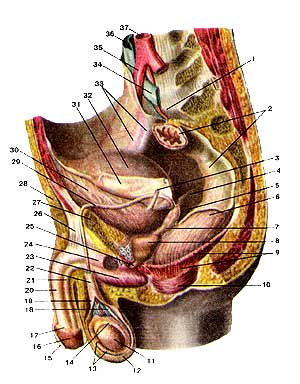 Мужские половые органы (organa genitalia masculine). Вид слева. Левые отделы стенок таза и левая стенка мошонки удалены: 1-мыс крестца; 2-прямая кишка (покрыта брюшиной); 3-мочеточник (левый); 4-прямокишечно-мочепузырная складка (правая); 5-прямокишечно-мочепузырное углубление; 6-прямая кишка (обнажена мышечная оболочка); 7-семенной пузырек (левый); 8-предстательная железа; 9-мышца, поднимающая задний проход; 10-наружный сфинктер заднего прохода; 11-яичко; 12-мошонка; 13-серозная (влагалищная) оболочка яичка; 14-придаток яичка; 15-крайняя плоть (полового члена); 16-головка полового члена; 17-венец головки; 18-семявыносящий проток; 19-внутренняя семенная фасция. 20-пещеристое тело полового члена; 21-губчатое тело половою члена; 22-семенной канатик; 23-луковица полового члена; 24-седалищно-пешеристая мышца; 25-перепончатая часть мужского мочеиспускательного канала (мужской уретры); 26-связка, поддерживашая половой член; 27-лобковая кость: 28-подкожная клетчатка; 29-семявыносящий проток; 30-мочевой пузырь (мышечная оболочка); 31-брюшина; 32-мочевой пузырь; покрытый брюшиной; 33-наружные подвздошные артерия и вена; 34-левая общая подвздошная вена; 35-правая общая подвздошная артерия; 36-нижняя полая вена; 37-аорта.
ЯИЧКИ. СТРОЕНИЕ ЯИЧЕК.
Яички, testes
Мужские половые органы (organa genitalia masculine). Вид слева. Левые отделы стенок таза и левая стенка мошонки удалены: 1-мыс крестца; 2-прямая кишка (покрыта брюшиной); 3-мочеточник (левый); 4-прямокишечно-мочепузырная складка (правая); 5-прямокишечно-мочепузырное углубление; 6-прямая кишка (обнажена мышечная оболочка); 7-семенной пузырек (левый); 8-предстательная железа; 9-мышца, поднимающая задний проход; 10-наружный сфинктер заднего прохода; 11-яичко; 12-мошонка; 13-серозная (влагалищная) оболочка яичка; 14-придаток яичка; 15-крайняя плоть (полового члена); 16-головка полового члена; 17-венец головки; 18-семявыносящий проток; 19-внутренняя семенная фасция. 20-пещеристое тело полового члена; 21-губчатое тело половою члена; 22-семенной канатик; 23-луковица полового члена; 24-седалищно-пешеристая мышца; 25-перепончатая часть мужского мочеиспускательного канала (мужской уретры); 26-связка, поддерживашая половой член; 27-лобковая кость: 28-подкожная клетчатка; 29-семявыносящий проток; 30-мочевой пузырь (мышечная оболочка); 31-брюшина; 32-мочевой пузырь; покрытый брюшиной; 33-наружные подвздошные артерия и вена; 34-левая общая подвздошная вена; 35-правая общая подвздошная артерия; 36-нижняя полая вена; 37-аорта.
ЯИЧКИ. СТРОЕНИЕ ЯИЧЕК.
Яички, testes (греч. — orchis s. didymis), представляют пару овальной формы несколько сплющенных с боков тел, расположенных в мошонке. Длинник яичка равен в среднем 4 см, поперечник — 3 см, масса от 15 до 25 г. В яичке различают две поверхности — facies medialis и lateralis, два края — margo anterior и posterior и два конца — extremitas superior и inferior.
При нормальном положении яичка в мошонке верхний конец его обращен вверх, кпереди и латерально, вследствие чего и нижний конец обращен не только книзу, но также кзади и медиально. Левое яичко обычно опущено несколько ниже, чем правое. К заднему краю яичка подходят семенной канатик, funiculus spermaticus, и придаток яичка, epididymis; последний располагается вдоль заднего края. Epididymis представляет собой узкое длинное тело, в котором различают верхнюю, несколько утолщенную часть — головку придатка, caput epididymitis, и нижний, более заостренный конец, cauda epididymidis; промежуточный участок составляет тело, corpus epididymidis. В области тела между передней вогнутой поверхностью придатка и яичком имеется пазуха, sinus epididymidis, выстланная серозной оболочкой и открытая в латеральную сторону.
На верхнем конце яичка нередко находится маленький отросток — appendix testis; на разрезе он состоит из тонких канальцев; представляет, по-видимому, рудиментарный остаюк ductus paramesonephricus. На головке придатка встречается appendix epididymidis, сидящий обычно на ножке (остаток mesonephros).
Строение яичка.
Яичко окружено плотной фиброзной оболочкой беловатой окраски, tunica albuginea, лежащей непосредственно на веществе или паренхиме яичка, parenchyma testis.
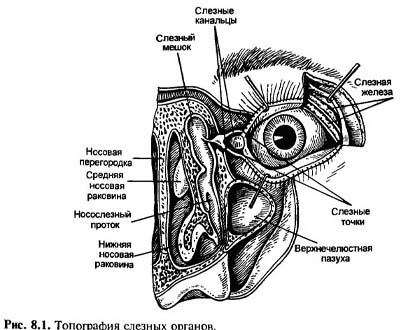
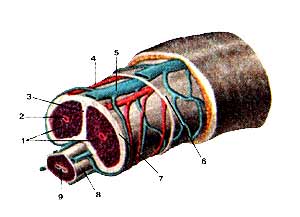
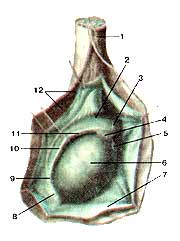 Яичко (testis) и придаток яичка (epididy-mus) Латеральная сторона. Оболочки яичка вскрыты: 1-семенной канатик; 2-головка придатка яичка; 3-принесок придатка яичка; 4-верхняя связка придатка яичка; 5-привесок яичка; 6-яичко; 7-влагалищная оболочка яичка; 8-нижняя связка придатка яичка; 9-хвост придатка яичка; 10-пазуха придатка яичка; 11-тело придатка яичка; 12-внутренняя семенная фасция.
Яичко (testis) и придаток яичка (epididy-mus) Латеральная сторона. Оболочки яичка вскрыты: 1-семенной канатик; 2-головка придатка яичка; 3-принесок придатка яичка; 4-верхняя связка придатка яичка; 5-привесок яичка; 6-яичко; 7-влагалищная оболочка яичка; 8-нижняя связка придатка яичка; 9-хвост придатка яичка; 10-пазуха придатка яичка; 11-тело придатка яичка; 12-внутренняя семенная фасция.
По заднему краю фиброзная ткань оболочки вдается на короткое расстояние внутрь железистой ткани яичка в виде неполной вертикальной перегородки или утолщения, носящего название mediastinum testis; от mediastinum лучеобразно отходят фиброзные перегородки — septula testis, которые своими наружными концами прикрепляются к внутренней поверхности tunica albuginea и, таким образом, делят всю паренхиму яичка на дольки, lobuli testis. Число долек яичка доходит до 250 —300. Вершины долек обращены к mediastinum, а основания — к tunica albuginea. Придаток яичка также имеет tunica albuginea, но более тонкую.
Паренхима яичка состоит из семенных канальцев, в которых различают два отдела — tubuli seminiferi contorti и tubuli seminiferi recti. В каждой дольке имеется 2 — 3 канальца и более. Имея извилистое направление в самой дольке, семенные канальцы, tubuli seminiferi contorti, приближаясь к mediastinum, соединяются друг с другом и непосредственно у mediastinum суживаются в короткие прямые трубки, tubuli seminiferi recti. Прямые канальцы открываются в сеть ходов — rete testis, расположенную в толще mediastinum. Из сети яичка открываются 12—15 выносящих канальцев — ductuli efferentes testis, которые направляются к головке придатка.
По выходе из яичка выносящие канальцы становятся извилистыми и образуют ряд конических долек придатка, lobuli s. coni epididymidis. Ductuli efferentes открываются в одиночный канал придатка, ductus epididymidis, который, образуя многочисленные изгибы, продолжается в ductus deferens. Будучи расправлен, канал придатка достигает 3 —4 м. Ductuli efferentes, lobuli epididymidis и начальный отдел канала придатка образуют в совокупности головку придатка. На придатке яичка встречаются отклоняющиеся протоки, ductuli aberrdntes. Тотчас выше головки придатка, кпереди от семенного канатика, встречается небольшое тело, paradidymis, которое представляет рудиментарный остаток первичной почки.
Местом образования спермиев, spermium, — основной части мужского семени являются лишь tubuli seminiferi contorti. Tubuli recti и канальцы сети яичка принадлежат уже к выводящим путям.
Жидкая составная часть
семени — sperma — только в незначительном количестве продуцируется яичками. Она представляет собой главным образом продукт выделения придаточных желез полового аппарата, открывающихся в выводящие пути.
Семявыносящий проток, ductus deferens
Семявыносящий проток, ductus deferens, парный, будучи непосредственным продолжением канала придатка, отличается от последнего большей толщиной своих сгенок. Отделенный от яичка сосудами (а. и v. testiculares), поднимается кверху и входит в состав семенного канатика. В последнем он располагается позади сосудов и легко прощупывается благодаря плотной консистенции своих стенок.
В составе семенного канатика он поднимается вертикально вверх к поверхностному паховому кольцу. Пройдя в паховом канале косо вверх и латерально, он у глубокого пахового кольца оставляет vasa testiculares (последние направляются в поясничную область) и идет вниз и назад по боковой стенке таза, будучи прикрыт брюшиной. Достигнув мочевого пузыря, проток загибается ко дну мочевого пузыря и подходит к предстательной железе. В нижнем своем отделе он заметно расширяется в виде ампулы семявыносящего протока, ampulla ductus deferentis.
Длина ductus deferens равняется 40 — 45 см. Средний диаметр 2,5 мм, ширина его просвета всего 0,2 — 0,5 мм. Стенка ductus deferens состоит из трех слоев: наружной фиброзной оболочки, tunica adventitia, затем средней мышечной, tunica muscularis, и внутренней слизистой, tunica mucosa.
Семенные пузырьки, vesiculae seminales.
Семенные пузырьки, vesiculae seminales, лежат латерально от семявыносящих протоков, между дном мочевого пузыря и прямой кишкой. Каждый семенной пузырек представляет собой сильно извитую трубку, имеющую в расправленном виде длину до 12 см, в нерасправленном — 5 см.
Нижний заостренный конец семенного пузырька переходит в узкий выделительный проток, ductus excretorius, который соединяется под острым углом с ductus deferens той же стороны, образуя вместе с ним семявыбрасывающий проток, ductus ejaculatorius. Последний представляет собой тоненький каналец, который, начавшись от места слияния ductus deferens и ductus excretorius, проходит через тощу предстательной железы и открывается в предстательную часть мочеиспускательного канала узким отверстием у основания семенного бугорка.
Длина семявыбрасывающего протока около 2 см. Стенки семенных пузырьков состоят из тех же слоев, что и ductus deferens. Семенные пузырьки представляют собой секреторные органы, которые вырабатывают жидкую часть семени.
Оболочки яичка и семенного канатика.
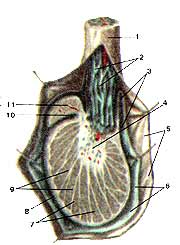
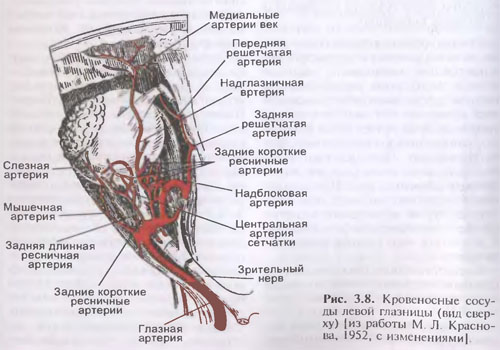
B]Яичко (testis) и придаток яичка (epididy-mus) на продольном разрезе[/B]
1-семенной канатик; 2-лозовидное сплетение 11 (венозное); 3-серозная 10 полость; 4-средостение яичка; 5-внутренняя семенная фасция; 6-влагалищная оболочка яичка; 7-дольки яичка; 8-белочная оболочка яичка; 9-перегородочки яичка; 10-пазуха придатка яичка; 11-головка придатка яичка.
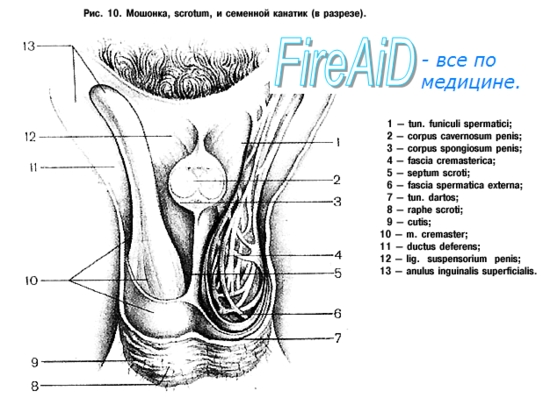
Оболочки яичка и семенного канатика, считая снаружи, следующие: кожа, tunica dartos, fascia spermatica externa, fascia cremasterica, m. cremaster, fascia spermatica interna, tunica vaginalis testis. Такое большое число оболочек яичка соответствует определенным слоям передней брюшной стенки. Яичко при своем смещении из брюшной полости как бы увлекает за собой брюшину и фасции мышц живота и оказывается окутанным ими.
[
1. Кожа мошонки тонкая и имеет более темную окраску по сравнению с другими участками тела. Она снабжена многочисленными крупными сальными железами, секрет которых имеет особый характерный запах.
2. Tunica dartos, мясистая оболочка, расположена тотчас под кожей. Она представляет собой продолжение подкожной соединительной ткани из паховой области и промежности, но лишена жира. В ней находится значительное количество гладкой мышечной ткани. Tunica dartos образует для каждого яичка по одному отдельному мешку, соединенному друг с другом по средней линии, так что получается перегородка, septum scroti, прикрепляющаяся по линии raphe.
3. Fascia spermatica externa — продолжение поверхностной фасции живота.
4. Fascia cremasterica представляет собой продолжение fascia intercruralis, отходящей от краев поверхностного пахового кольца; она покрывает m. cremaster, а поэтому и называется fascia cremasterica.
5. М. cremaster состоит из пучков исчерченных мышечных волокон, являющихся продолженем m. transversus abdominis (см. «Миология»). При сокращении т. cremaster яичко подтягивается кверху.
6. Fascia spermatica interna, внутренняя семенная фасция, расположена тотчас под m. cremaster. Она представляет собой продолжение fascia transversalis, охватывает кругом все составные части семенного канатика и в области яичка прилежит к наружной поверхности его серозного покрова.
7. Tunica vaginalis testis, влагалищная оболочка яичка, происходит за счет processus vaginalis брюшины и образует замкнутый серозный мешок, состоящий из двух пластинок: lamina parietalis — пристеночная пластинка и lamina visceralis — висцеральная пластинка. Висцеральная пластинка тесно срастается с белочной оболочкой яичка и переходит также на придаток. Между латеральной поверхностью яичка и средней частью придатка (телом) висцеральная пластинка заходит в щеле-видное пространство между ними, образуя sinus epididymidis.
Вдоль заднего края яичка по месту выхода сосудов висцеральная пластинка переходит в париетальную. Между обращенными друг к другу поверхностями париетальной и висцеральной пластинок имеется щелевидное-пространство — cavum vaginale, в котором в патологических случаях может скопиться большое количество серозной жидкости и обусловить водянку яичка.
Половой член, penis.
Половой член, penis, составляет вместе с мошонкой наружные половые органы. В его состав входят три тела: парное пещеристое, corpus cavernosum penis, и непарное губчатое, corpus spongiosum penis. Название этих тел обусловлено тем, что они состоят из многочисленных перекладин, фиброзно-эластических тяжей с примесью неисчерченных мышечных волокон, среди густого сплетения которых есть промежутки, пещеры, выстланные эндотелием и заполненные кровью.
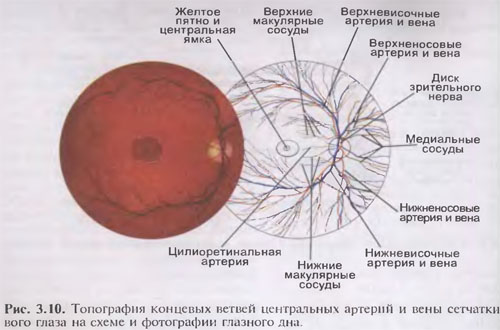
 Мужские половые органы (organa genitalia masculine). Предстательная желе:и (prostata), пешеристые тела (corpora cav-crnosa), губчатое тело (corpus spongiosus) полового члена, бульооуретральные железы (glandulae hulbouretralis) Мочевой пузырь и мочеиспускательный канал вскрыты: 1-мочевой пузырь; 2-мочеточник; 3-треугольник мочевого пузыря; 4-язычок пузыря; 5-внутреннее отверстие мочеиспускательного канала; 6-предстательная железа; 7-предстательная часть (мочеиспускательного канала); 8-перепончатая часть (мочеиспускательного канала); 9-бульбоуретральная железа; 10-луковица полового члена; 11-пешеристое тело полового члена; 13-лакуны мочеиспускательного канала; 14-головка полового члена; 15-крайняя плоть; 16-наружное отверстие мочеиспускательного канала; 17-ладьевидная ямка мочеиспускательного канала; 18-губчатое тело полового члена; 19-протоки бульбоуретральных желез; 20-семенной холмик; 21-отверстие правого мочеточника; 22-складки слизистой оболочки мочевого пузыря.
Мужские половые органы (organa genitalia masculine). Предстательная желе:и (prostata), пешеристые тела (corpora cav-crnosa), губчатое тело (corpus spongiosus) полового члена, бульооуретральные железы (glandulae hulbouretralis) Мочевой пузырь и мочеиспускательный канал вскрыты: 1-мочевой пузырь; 2-мочеточник; 3-треугольник мочевого пузыря; 4-язычок пузыря; 5-внутреннее отверстие мочеиспускательного канала; 6-предстательная железа; 7-предстательная часть (мочеиспускательного канала); 8-перепончатая часть (мочеиспускательного канала); 9-бульбоуретральная железа; 10-луковица полового члена; 11-пешеристое тело полового члена; 13-лакуны мочеиспускательного канала; 14-головка полового члена; 15-крайняя плоть; 16-наружное отверстие мочеиспускательного канала; 17-ладьевидная ямка мочеиспускательного канала; 18-губчатое тело полового члена; 19-протоки бульбоуретральных желез; 20-семенной холмик; 21-отверстие правого мочеточника; 22-складки слизистой оболочки мочевого пузыря.
Corpora cavernosa penis представляет собой два длинных цилиндрических тела с заостренными концами, из которых задние расходятся и образуют crura penis, прикрепляющиеся к нижним ветвям лобковых костей. Эти два тела покрыты общей белочной оболочкой, tunica albuginea corporum cavernosorum, которая в промежутке между ними образует septum penis. Соответственно перегородке на верхней поверхности находится борозда для v. dorsalis penis, а на нижней поверхности — для corpus spongiosum penis.
Corpus spongiosum penis, покрытое tunica albuginea corporis spongiosi, лежит снизу пещеристых тел члена и пронизано во всю длину мочеиспускательным каналом. Оно имеет меньший, чем два других пещеристых тела, диаметр (1 см), но в отличие от них утолщается на обоих концах, образуя спереди головку члена, glans penis, а сзади луковицу — bulbus penis.
Задняя часть полового члена, прикрепленная к лобковым костям, носит название корня, radix penis. Кпереди половой член оканчивается головкой, glans penis. Промежуточная между головкой и корнем часть называется телом, corpus penis. Верхняя поверхность тела шире нижней и носит название спинки, dorsum penis. К нижней поверхности прилежит corpus spongiosum penis. На головке члена имеется вертикальная щель — наружное отверстие мочеиспускательного канала, ostium urethrae externum; головка с дорсальной и с латеральной сторон несколько выдается над уровнем пещеристых тел; этот край головки носит название corona glandis, а сужение позади него — collum gldndis.
 Пещеристые и губчатое тела (corpora cavernosa et corpu spongiosus) полового члена Губчатое тело частично отделено от пещеристых тел: 1-наружное отверстие мочеиспускательного канала; 2-головка полового члена; 3-венец головки; 4-губчатое тело полового члена; 5-луковично-пешеристая мышца; 6-луковица полового члена; 7-бульбоуретральная железа; 8-седалишный бугор; 9-глубокая поперечная мышца промежности;10-нижняя фасция мочеполовой диафрагмы; 11-наружная мышца, сжимающая мочеиспускательный канал (наружный сфинктер мочеиспускательного канала); 12-ножка полового члена; 13-седалищно-пещеристая мышца; 14-глубокая фасция полового члена; 15-пещеристые тела полового члена.
Пещеристые и губчатое тела (corpora cavernosa et corpu spongiosus) полового члена Губчатое тело частично отделено от пещеристых тел: 1-наружное отверстие мочеиспускательного канала; 2-головка полового члена; 3-венец головки; 4-губчатое тело полового члена; 5-луковично-пешеристая мышца; 6-луковица полового члена; 7-бульбоуретральная железа; 8-седалишный бугор; 9-глубокая поперечная мышца промежности;10-нижняя фасция мочеполовой диафрагмы; 11-наружная мышца, сжимающая мочеиспускательный канал (наружный сфинктер мочеиспускательного канала); 12-ножка полового члена; 13-седалищно-пещеристая мышца; 14-глубокая фасция полового члена; 15-пещеристые тела полового члена.
Кожа полового члена у основания головки образует свободную складку, которая носит название крайней плоти, preputium. На нижней стороне головки члена крайняя плоть соединена с кожей головки уздечкой, frenulum preputii. Вокруг corona glandis и на внутреннем Листке крайней плоти расположены различной величины сальные железки, glandulae preputiales. Секрет этих желез входит в состав препуциальной смазки, smegma preputii, собирающейся в желобе между glans penis и preputium. Между головкой и крайней плотью остается пространство — полость крайней плоти, открывающееся спереди отверстием, которое пропускает головку при отодвигании крайней плоти кзади. На нижней поверхности члена, по средней линии от frenulum preputii, внизу заметен шов, raphe, указывающий место сращения первоначально двух отдельных половин. С полового члена шов простирается кзади на мошонку и промежность. Три тела полового члена соединяются в одно целое окружающей их fascia penis, лежащей под рыхлой подкожной клетчаткой. Кроме того, корень члена укрепляется связками.
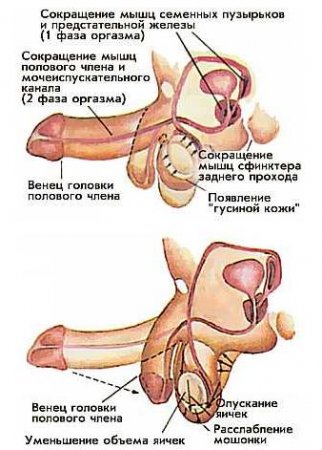
Величина penis изменяется в зависимости от количества крови в камерах пещеристых и губчатого тел. Кровь приносится к половому члену через аа. profundae et dorsalis penis. Артериальные ветви, проходя в соединительнотканных перегородках, распадаются на тонкие завитковые артерии, которые открываются прямо в кавернозные пространства. Отводящие кровь вены, venae cavernosae, начинаются частью в центральных участках пещеристых тел, частью более периферически и вливаются в vv. profundae penis и в v. dorsalis penis. Благодаря особому устройству кровеносных сосудов члена кровь в пещеристых телах может задерживаться, что приводит к их уплотнению при эрекции.
 Сосуды ( кровоснабжение ) полового члена. Нервы ( иннервация ) полового члена.
Сосуды ( кровоснабжение ) полового члена. Нервы ( иннервация ) полового члена.
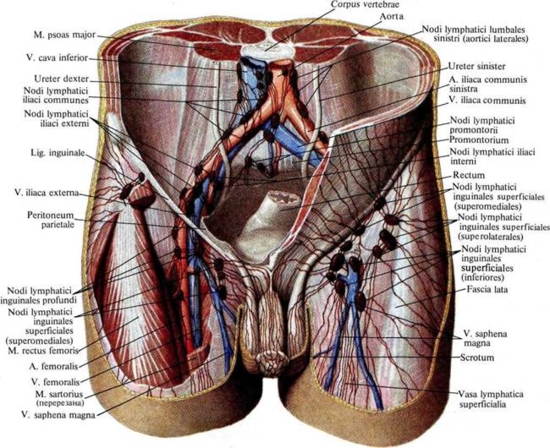
Сосуды и нервы: артерии полового члена являются ветвями a. femoralis (аа. pudendae externae) и a. pudenda interna. Венозный отток происходит по vv. dorsales penis superficialis et profundae в v. femoralis и в plexus venosus vesicalis. Лимфоотток осуществляется в nodi lymphatici inguinales и узлы полости малого таза.
Кроме анатомического деления мочеиспускательного канала на 3 части, в урологической клинике (соответственно течению воспалительных процессов) различают 2 отдела его: переднюю уретру, т.е. pars spongiosa, и заднюю — остальные две части. Границей между ними служит m. sphincter urethrae, который препятствует проникновению инфекции из передней уретры в заднюю.
На всем протяжении слизистой оболочки, за исключением ближайшего к наружному отверстию участка, в канал открываются многочисленные железки — glandulae urethrales. Кроме того, преимущественно на верхней стенке мочеиспускательного канала, в особенности кпереди от луковицы, находятся углубления — lacunae urethrales; отверстия их обращены кпереди и прикрыты клапанообразными заслонками. Кнаружи от под-слизистой основы располагается слой неисчерченных мышечных волокон (изнутри продольные, снаружи циркулярные).
 Мочеиспускательный канал
Мочеиспускательный канал на своем пути имеет S-образную изогнутость. При поднимании кверху pars spongiosa передняя кривизна выпрямляется и остается один изгиб с вогнутостью, обращенной к symphysis pubica. Большая фиксированность задней кривизны обеспечивается ligg. puboprosta-tica, идущими от симфиза к предстательной железе, diaphragma urogenitale (через нее проходит pars membranacea urethrae), а также lig. suspen-sorium penis, соединияющей penis с симфизом.
Калибр просвета мочеиспускательного канала не везде одинаков. Измерение металлических слепков дало такие цифры: место соединения pars spongiosa и pars membranacea — 4,5 мм, наружное отверстие — 5,7 мм, середина pars prostatica — 11,3 мм, в области bulbus — 16,8 мм. Возможно, что семя перед выбрасыванием предварительно собирается в расширенной соответственно bulbus части канала. У взрослого можно считать максимальным для введения в канал катетер диаметром 10 мм.
Афферентная иннервация проводится по n. pudendus, эфферентная симпатическая — из plexus hypogastrics inferior, парасимпатическая — nn. erigentes.
 Пещеристые и губчатое тела (corpora cavernosa et corpi spongiosus) полового члена и мужской мочеиспускательный канал (urethra masculina) на поперечном разрезе 1-пещеристые тела; 2-глубокая артерия полового члена; 3-перегородка полового члена;
1. Pars prostatica, предстательная часть,
Пещеристые и губчатое тела (corpora cavernosa et corpi spongiosus) полового члена и мужской мочеиспускательный канал (urethra masculina) на поперечном разрезе 1-пещеристые тела; 2-глубокая артерия полового члена; 3-перегородка полового члена;
1. Pars prostatica, предстательная часть, ближайшая к мочевому пузырю, проходит через предстательную железу. Длина этого отдела около 2,5 см. Предстательная часть, особенно ее средний отдел, является наиболее широким и растяжимым участком мочеиспускательного канала. На задней стенке находится небольшое срединное возвышение — colliculus seminalis, семенной бугорок около 1,5 см длиной. На верхушке семенного бугорка щелевидное отверстие ведет в небольшой слепой кармашек, расположенный в толще предстательной железы, который носит название utriculus prostaticus (предстательная маточка). Название указывает на происхождение этого образования из слившихся нижних концов ductus paramesonephricus, из которых у женщины развиваются матка и влагалище. По сторонам от входа в utriculus prostaticus находятся на colliculus seminalis маленькие отверстия семявыбрасывающих протоков (по одному справа и слева).
Латерально от семенного бугорка по обеим сторонам открываются многочисленные отверстия простатических железок. По окружности предстательной части мочеиспускательного канала имеется, кольцо мышечных волокон, составляющих часть гладкой мышечной ткани предстательной железы, усиливающих сфинктер мочевого пузыря, sphincter vesicae (гладкомышечный, непроизвольный).
2. Pars membranacea, перепончатая часть , представляет собой участок мочеиспускательного канала на протяжении от верхушки предстательной железы до bulbus penis; длина ее около 1 см. Таким образом, этот отдел канала является наиболее коротким и в то же время наиболее узким из всех трех. Он лежит кзади и книзу от lig. arcuatum pubis, прободая на своем пути diaphragma urogenitale с ее верхней и нижней фасциями; нижний конец перепончатой части на месте прободения нижней фасции представляет собой самый узкий и наименее растяжимый участок канала, что необходимо учитывать при введении катетера, чтобы не прорвать канал. Перепончатая часть мочеиспускательного канала окружена мышечными пучками произвольного сфинктера, m. sphincter utethrae.
3. Pars spongiosa, губчатая часть, длиной около 15 см, окружена тканью corpus spongiosum penis. Часть канала соответственно bulbus несколько расширена; на остальном протяжении до головки диаметр канала равномерный, в головке на протяжении приблизительно 1 см канал опять расширяется, образуя ладьевидную ямку, fossa naviculdris urethrae. Наружное отверстие является малорастяжимой частью мочеиспускательного канала, что следует учитывать при вставлении зонда.
Мужской мочеиспускательный канал.
Мужской мочеиспускательный канал, urethra masculina, представляет трубку около 18 см длиной, простирающуюся от мочевого пузыря до наружного отверстия мочеиспускательного канала, ostium urethrae externum, на головке полового члена. Urethra служит не только для выведения мочи, но также для прохождения семени, которое поступает в мочеиспускательный канал через ductus ejaculatorius. Мочеиспускательный канал проходит через различные образования, поэтому в нем различают три части: pars prostatica, pars membranacea и pars spongiosa.
Сосуды ( кровоснабжение ) и нервы ( иннервация ) мочеиспускательного канала. Акт мочеиспускания.
Артерии мочеиспускательного канала происходит из ветвей a. pudenda interna. Разные отделы канала питаются из различных источников: pars prostatica — из ветвей а. гесtalis media и a. vesicalis inferior; pars membranacea — из a. rectalis inferior и a. perinealis; pars spongiosa — из a. pudenda interna. В васкуляризации стенок канала участвуют также a. dorsalis penis и a. profunda penis. Венозная кровь оттекает к венам penis и к венам мочевого пузыря. Лимфоотток происходит из pars prostatica к лимфатическим сосудам prostatae, из pars membranacea и pars spongiosa — к паховым узлам. Иннервация осуществляется из nn. perinei и п. dorsalis penis (из п. pudendus), а также из вегетативного сплетения, plexus prostaticus.
Акт мочеиспускания осуществляется следующим образом: сокращающийся m. detrusor urinae выжимает мочу из мочевого пузыря, которая поступает в мочеиспускательный канал, открывающийся благодаря расслаблению своих сфинктеров: непроизвольного (m. sphincter vesicae) и произвольного (т. sphincter urethrae).
У мужчин происходит также расслабление мышечной части предстательной железы, выполняющей функции третьего (непроизвольного) сфинктера. Закрытие мочевого пузыря происходит при расслаблении m. detrusor и сокращении названных сфинктеров.
В последнее время появились сведения о наличии четвертого сфинктера, расположенного ниже предстательной железы. Имеется и другой взгляд, согласно которому сфинктер мочевого пузыря не существует, а его роль выполняет у женщин вся уретра, а у мужчин — пречстательная и перепончатая части мочеиспускательного канала, а также эластическая ткань, заложенная в стенках уретры.
Бульбоуретральные железы. Предстательная железа. Анатомия простаты ( предстательной железы ).
Бульбоуретральные железы. Glandulae bulbourethrales представляют собой две железки величиной каждая с горошину, которые располагаются в толще diaphragma urogenitale над задним концом bulbus penis, кзади от pars membranacea urethrae. Выводной проток этих желез открывается в губчатую часть мочеиспускательного канала в области bulbus. Железы выделяют тягучую жидкость, которая защищает стенки мочеиспускательного канала от раздражения мочой.
Артерии к бульбоуретральным железам подходят из a. pudenda interna. Венозный отток происходит в вены bulbus и diaphragma urogenitale. Лимфатические сосуды идут к nodi lymphatici iliaci interni.
Иннервируются железы из п. pudendus.
Предстательная железа. Анатомия простаты ( предстательной железы ).
Предстательная железа, prostata (prostates, греч. от proistanai — представить, выдаваться вперед), представляет собой меньшей частью железистый, большей частью мышечный орган, охватывающий начальную часть мужского мочеиспускательного канала. Как железа она выделяет секрет, составляющий важную часть спермы и стимулирующий спермин, и потому развивается ко времени полового созревания. Имеются указания и на наличие эндокринной функции железы.
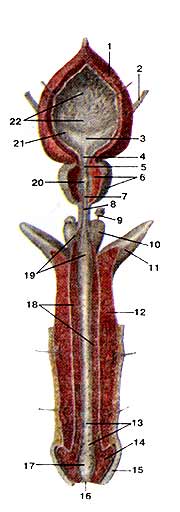
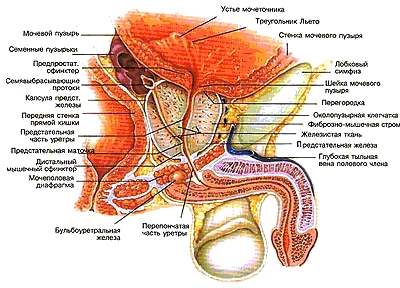
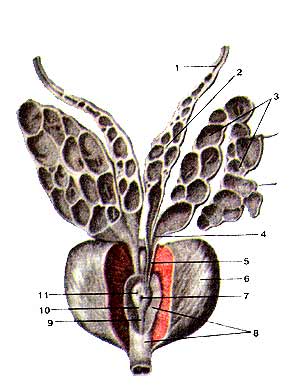 Предстательная железа (prostata), семенные пузырьки (vesiculae seminales) и семявыносяшие протоки (ductus deierentis) вскрыты Простатическая часть мочеиспускательною канала частично вскрыта: 1-семявыносящий проток; 2-ампула семявыносящею протока; 3-семенной пузырек; 4-выделительный проток семенною пузырька; 5-семявыбрасываюший проток; 6-предстательная железа; 7-мужская маточка; 8-мочеиспускательный канал; 9-гребень мочеиспускательного канала; 10-семенной холмик; 11-устье семявыбрасываюшего протока.
Предстательная железа (prostata), семенные пузырьки (vesiculae seminales) и семявыносяшие протоки (ductus deierentis) вскрыты Простатическая часть мочеиспускательною канала частично вскрыта: 1-семявыносящий проток; 2-ампула семявыносящею протока; 3-семенной пузырек; 4-выделительный проток семенною пузырька; 5-семявыбрасываюший проток; 6-предстательная железа; 7-мужская маточка; 8-мочеиспускательный канал; 9-гребень мочеиспускательного канала; 10-семенной холмик; 11-устье семявыбрасываюшего протока.
Как мышца она является непроизвольным сфинктером мочеиспускательного канала, в частности препятствующим истечению мочи во время эякуляции, вследствие чего моча и сперма не смешиваются. До наступления половой зрелости является исключительно мышечным органом, а ко времени полового созревания (17 лет) становится и железой. Формой и величиной prostata напоминает каштан. В ней различают основание, basis prostatae, обращенное к мочевому пузырю, и верхушку, apex, примыкающую к diaphragma urogenitale.
Передняя выпуклая поверхность железы, facies anterior, , обращена к лобковому симфизу, от которого отделяется рыхлой клетчаткой и заложенным в ней венозным сплетением (plexus prostaticus); поверх этого сплетения лежат ligg. pubovesicalia. Задняя поверхность прилежит к прямой кишке, отделяясь от последней только пластинкой тазовой фасции (septum rectovesicale); поэтому ее можно прощупать у живого на передней стенке прямой кишки пальцем, введенным per rectum. Urethra проходит через предстательную железу от ее основания к верхушке, располагаясь в срединной плоскости, ближе к передней поверхности железы, чем к задней.
Семявыбрасывающие протоки входят в железу на задней поверхности, направляются в толще ее вниз, медиально и кпереди и открываются в pars prostatica urethrae. Участок железы, расположенный между обоими ductus ejaculatorii и задней поверхностью urethrae, имеющий клиновидную форму, составляет средний отдел железы, isthmus prostatae (lobus medius). Остальную, большую, часть составляют lobi dexter et sinister, которые, однако, с поверхности не разграничены резко друг от друга.
Средняя доля представляет значительный хирургический интерес, так как, увеличиваясь при гипертрофии предстательной железы, может быть причиной расстройства мочеиспускания.
Наибольшим диаметром предстательной железы является поперечный (близ основания); он равен в среднем 3,5 см, переднезадний — 2 см, вертикальный — 3 см.
Prostata окружена фасциальными листками, происходящими за счет fascia pelvis и образующими вместилище, в котором находится венозное сплетение, plexus prostaticus.
Кнутри от фасциальный оболочки находится capsula prostatica, состоящая из гладкой мышечной и соединительной ткани. Ткань prostatae состоит из желез (parenchyma glandularae), погруженных в основу, состоящую главным образом из мышечной ткани, substantia muscularis; дольки ее состоят из тонких, слегка разветвленных трубочек, впадающих в ductuli prostatici (числом около 20 — 30), которые открываются на задней стенке предстательной части urethrae по сторонам от colliculus seminalis. Часть предстательной железы кпереди от проходящего через нее мочеиспускательного канала состоит почти исключительно из мышечной ткани.
Сосуды и нервы: prostata получает питание из аа. vesicales inferiores и аа. rectales mediae. Вены вступают в plexus vesicalis et prostaticus, из которого выносят кровь vv. vesicales inferiores; сосуды предстательной железы достигают полного развития лишь после наступления половой зрелости.
Лимфатические сосуды вливаются в узлы, расположенные в передних отделах полости таза.
Нервы происходят из plexus hypogastrics inf.
Пути выведения семени в последовательном порядке: tubuli seminiferi recti, rete testis, ductuli efferentes, ductus epididymidis, ductus deferens, ductus ejaculatorius, pars prostatica urethrae и остальные части мочеиспускательного канала.
с использованием материалов книги Григория Белоголовского «АНАТОМИЯ ЧЕЛОВЕКА»